亂拞惈巕彫妏嶶棎偑娤傞僙儖儘乕僗偺峺慺怗攠廳崌偲慻怐宍惉亃
亂拞惈巕彫妏嶶棎偑娤傞僙儖儘乕僗偺峺慺怗攠廳崌偲慻怐宍惉亃
仠愭抂婎慴尋媶僙儞僞乕 僜僼僩儅僞乕拞惈巕嶶棎尋媶Gr 揷拞 岹榓 丒 彫愹 抭 丒 嫶杮 抾帯
僙儖儘乕僗偼抧媴忋偵嵟傕懡偔懚嵼偡傞崅暘巕偱偁傝丄変乆偺擔乆偺惗妶偵晄壜寚側慺嵽偱偁傞丅捠忢僙儖儘乕僗偼怉暔傗旝惗暔偵傛傞揤慠崌惉偵傛偭偰惗惉偝傟偰偄傞丅
挿偄崅暘巕尋媶偺楌巎偺拞偱僙儖儘乕僗偺恖岺崌惉偼廳梫側壽戣偲偝傟側偑傜丄惉岟偵偼帄傜側偐偭偨丅偟偐偟1991擭彫椦傜偼丄
杮棃僙儖儘乕僗偺暘夝峺慺偱偁傞僙儖儔乕僛傪棙梡偟偨峺慺怗攠廳崌偵傛傝弶傔偰恖岺崌惉偵惉岟偟偨丅偨偩偟摼傜傟傞暘巕偺挿偝偼丄廳崌搙20掱搙偲嬌傔偰抁偄丅
峺慺怗攠偼摿掕偺婎幙偩偗偵嶌梡偟丄偐偮摿掕偺斀墳偺傒懀恑偡傞偲偄偆嬌傔偰桪傟偨慖戰惈傪寭偹旛偊偰偄傞丅偙偺傛偆側峺慺偺棙揰傪嵟戝尷妶梡偡傞偙偲偵傛偭偰丄
廬棃崌惉偑崲擄偱偁偭偨暋嶨側峔憿偺壔妛暔幙偺崌惉偑壜擻偲側傞偺偱偁傞丅
峺慺怗攠偵傛傞壔妛斀墳偱偼丄婎幙偑峺慺偺妶惈晹埵傪擣幆偡傞夁掱偑廳梫偱偁傞丅僙儖儘乕僗偺恖岺崌惉偺応崌偼丄
僋儗僼僩偲屇偽傟傞僞儞僷僋幙暘巕偺峚偑妶惈晹埵偵偁偨傞丅傑偨妶惈晹埵偐傜偼僙儖儘乕僗偑攔弌偝傟偦偺嬤朤偱寢徎壔偡傞丅
偮傑傝峺慺妶惈晹埵傗斀墳惗惉暔偺峔憿偑斀墳惈傪戝偒偔嵍塃偡傞壜擻惈偑偁傞丅
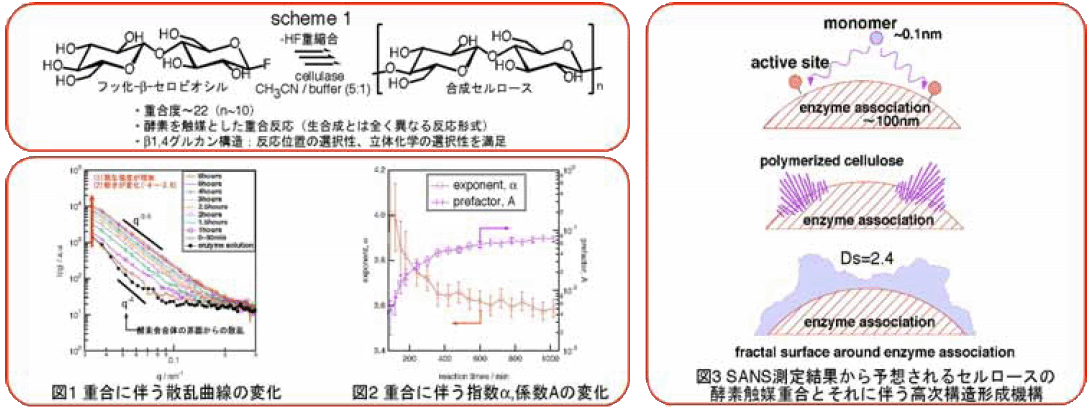
僜僼僩儅僞乕拞惈巕嶶棎尋媶僌儖乕僾偱偼峺慺怗攠廳崌偵傛傞僙儖儘乕僗偺恖岺崌惉傪拞惈巕彫妏嶶棎乮埲壓SANS乯偱偦偺応娤嶡偡傞偙偲傪帋傒偨丅偦偺寢壥丄
僫僲儊乕僩儖偐傜儅僀僋儘儊乕僩儖偵媦傇峀偄嬻娫僗働乕儖偱恖岺崌惉夁掱偵惗惉偝傟偨僙儖儘乕僗偑峺慺昞柺偱宍惉偡傞帺屓慻怐壔峔憿傪柧傜偐偵偡傞偙偲偑偱偒偨丅
拞惈巕嶶棎偼杮宯偺傛偆側乽壔妛斀墳偲峔憿宍惉偑枾愙偵娭楢偡傞宯乿偵偍偗傞峔憿娤嶡庤抜偲偟偰旕忢偵桪傟偰偄傞丅
幚尡偺徻嵶偼埲壓偺捠傝偱偁傞丅斀墳婎幙偲偟偰僼僢壔-β-僙儘價僆僔儖傪傾僙僩僯僩儕儖-恷巁娚徴梟塼(pH=5.0)(5/1, v/v)偵梟偐偟丄
偦偙偵峺慺偱偁傞僙儖儔乕僛傪廳崌怗攠偲偟偰壛偊傞 (Scheme 1)丅偙傟傪岤偝2mm偺僋僅乕僣僙儖偵擖傟丄30亷丄忢埑壓丄拞惈巕嶶棎偺應掕傪峴偭偨丅
梡偄偨憰抲偼JRR-3偵愝抲偝傟偨拞惈巕彫妏嶶棎憰抲SANS-J偍傛傃PNO偱偁傞丅SANS應掕寢壥傪恾1偵帵偡丅恾1傛傝帪娫偲偲傕偵嶶棎嫮搙I(q)偑扨挷偵憹壛偟偰偄傞丅
偙傟偼惗惉偟偨僙儖儘乕僗偑怗攠昞柺偱寢徎壔偟偨寢壥惗偠偨帺屓慻怐壔峔憿偺帪娫曄壔傪帵偡丅師偵嶶棎僨乕僞偺徻嵶傪傒偰傒傛偆丅
帪娫偵傛傜偢嶶棎嫮搙I(q)偲攇悢q偺椉懳悢僾儘僢僩偑捈慄忬偱偁傞偙偲偑暘偐傞丅偙偺孹偒偼偝傜偵2寘彫偝側攇悢傑偱懕偄偰偄傞丅
偙偙偱捈慄偺孹偒傪-a丄學悢A(斾奅柺愊)丄掕悢B(旕姳徛惈嶶棎)偲偟偰丄嶶棎嫮搙I(q)傪
I(q) = Aq-a + B丒丒丒(1)
偲嬤帡偟偰傒傞丅(1)幃傪梡偄偰奺廳崌帪娫偵偍偗傞嶶棎嬋慄傪僼傿僢僥傿儞僌偟偨丅偦偺寢壥摼傜傟偨a媦傃A偺帪娫曄壔傪恾2偵帵偡丅
恾2傛傝a偼帪娫偲偲傕偵尭彮偟A偼帪娫偲偲傕偵憹壛偡傞偙偲偑暘偐傞丅傑偨偄偢傟偺抣傕廳崌奐巒偐傜栺200暘偺娫偵媫寖偵曄壔偟丄埲崀傎傏堦掕抣偵嬤偯偄偰偄傞偙偲偑暘偐傞丅
偙傟偼儌僲儅乕徚旓検偺帪娫曄壔偲傕堦抳偟偰偄傞丅堦斒揑偵丄I(q)偲q偺娫偵(1)幃偺傛偆側儀僉忔懃偑惉傝棫偮偲偒丄峔憿偑帺屓憡帡惈(僼儔僋僞儖惈)傪傕偮偙偲偑帵嵈偝傟傞丅
崱偺応崌偼奅柺峔憿偵僼儔僋僞儖惈偑偁傞丅偙偙偱奅柺僼儔僋僞儖師尦傪Ds偲偡傞偲丄a偼
a = 6 - Ds (2 < Ds < 3)
偲昞偝傟傞丅偮傑傝僙儖儘乕僗偑峺慺昞柺偱宍惉偡傞寢徎峔憿偺奅柺(昞柺)偼廳崌偑恑傓偵偮傟偰師尦Ds=2.4傊偲曄壔偡傞偙偲偑暘偐傞丅 偙偺忬嫷傪柾幃揑偵帵偟偨偺偑恾3偱偁傞丅廳崌慜偵偼峺慺昞柺偼妸傜偐(師尦Ds=2丄偙傟偼億儘僢僪懃偲堦抳偡傞)偱偁偭偨偑丄 僙儖儘乕僗偺惗惉偵廬偭偰慹偄奅柺(2 < Ds < 3)傊偲曄壔偡傞條巕偑柧傜偐偲側偭偨丅偮傑傝丄傑偢峺慺夛崌懱偺妶惈晹埵偑儌僲儅乕傪擣幆偡傞偙偲偱廳崌偑奐巒偡傞丅 惗惉偝傟偨僙儖儘乕僗偼峺慺偐傜梀棧偡傞偙偲側偔偦偺昞柺嬤朤偵棷傑偭偰丄怴偨偵惗惉偝傟傞僙儖儘乕僗偲偲傕偵師尦Ds=2.4傪傕偭偨寢徎峔憿傪峺慺昞柺偺妶惈晹埵廃曈偵宍惉偡傞丅 怉暔傗旝惗暔偵傛傞揤慠崌惉偱惗惉偝傟傞僙儖儘乕僗偼廳崌搙悢愮偐傜悢枩偱偁傝恖岺崌惉偲偼斾妑偵側傜側偄傎偳戝偒偄丅 偙偺嵎偺梫場偲偟偰忋弎偟偨峺慺昞柺偱偺僙儖儘乕僗偺寢徎壔偑嫇偘傜傟傞丅偮傑傝寢徎壔偍傛傃寢徎峔憿偑廳崌斀墳偵塭嬁傪梌偊偰偄傞偺偱偼側偄偐丅 偙偺揰傪拞惈巕彫妏嶶棎偵傛偭偰柧傜偐偵偟丄偦傟偑廳崌搙偺惂屼偵宷偑傟偽慺惏傜偟偄偲峫偊丄峏側傞幚尡傪婇夋偟偰偄傞丅